In the Soviet Union in the mid-1950s adaptogens were defined by Nikolai Vasilievich Lazarev as harmless substances that can increase resistance to non-specific stress. Adaptogenic herbs are known as either ‘rasayana’ or ‘Qi tonic’ herbs in Ayurvedic and Chinese medicine, respectively. Although every adaptogen is often very different from one another and suited to different people, they are used by herbalists to treat a wide array of disorders, including fatigue, stress, immune, sexual, or nervous system problems, and depression (Winston, 2007). A popular current theory that may explain their broad applicability is that these herbs primarily modulate the sympatho-adrenal or hypothalamic-pituitary-adrenal axes of the endocrine system to adapt the organism to dealing with short or long-term stress (Panossian, 2017). Hormones such as cortisol are modulated in level by adaptogens and have receptors on both immune and nerve cells.
Recent molecular studies demonstrate adaptogen treatment can be considered to have a vaccination type effect on these stress response system. For example, it has been shown in C. elegans that adaptogens can induce translocation of the DAF-16 transcription factor from the cytoplasm into the nucleus, favouring an increased synthesis of proteins involved in stress resistance (Wiegant et al., 2009). Research on the ADAPT-252 compound (rhodiola, schisandra, eleutherococcus) shows that adaptogens can increase the synthesis of the heat shock protein HSP70 in vivo, a mediator of the adaptive stress response (Hernández‐Santana et al., 2014). The compromised ability of neurons to express HSP70 is associated with aging-related neurodegeneration and HSP70 application has been found to extend lifespan of animals and improve memory. This article describes in more detail the biology, history, and science surrounding the adaptogenic herbs. First we will need to delve into some terminology.
Figure 1. The adaptogen herbs eleutherococcus, schisandra, and rhodiola (from left to right).
Core terminology
Endocrine system: The endocrine system is the series of glands that release hormones into the blood to be carried to distant target organs. The major endocrine glands include; the pineal gland, the pituitary gland, pancreas, ovaries, testes, the thyroid gland, the parathyroid gland, and the adrenal glands.
The adrenal cortex: The adrenal cortex is the outer part of the adrenal gland and produces hormones such as cortisol and aldosterone.
Hypothalamus: The hypothalamus is a region of the brain containing small nuclei with a variety of functions, one of the most important of these is to link the nervous system with the endocrine system via the pituitary gland.
Pituitary: The pituitary or ‘master’ gland is a pea-sized structure found at the base of the brain immediately below the hypothalamus, to which it is attached via nerve fibres. It produces important hormones such as adrenocorticotropic hormone, growth hormone, thyroid stimulating hormone.
Hypothalamic-pituitary-adrenal (HPA) axis: The HPA axis comprises of 3 organs and their direct interactions, namely, the hypothalamus, the pituitary gland, and the adrenal glands. It is a major neuroendocrine system that controls reactions to stress and regulates many processes, including digestion, the immune system, mood and emotions, sexuality, and energy storage and expenditure.
Sympatho-adrenal-system (SAS): The SAS is a physiological connection between the sympathetic nervous system and the adrenal medulla and is important in the bodies response to outside stimuli. The SAS plays an important role in maintaining glucose levels, blood pressure, and other metabolic pathways that couple with bodily responses to the outside environment.
Cortisol: Cortisol is a steroid hormone that is produced by the adrenal glands which sit on top of each kidney. When released into the bloodstream it then acts on many different parts of the body and helps the body respond to stress, increases metabolism of glucose, controls blood pressure, and reduce inflammation.
Adrenocorticotrophic hormone-releasing factor (CRF): CRF is released by the paraventricular nucleus of the hypothalamus in response to stress. Its primary function is the stimulation of the pituitary synthesis of ACTH, as part of the HPA axis.
Vasopressin (AVP): AVP is a hormone synthesized in the hypothalamus and is released into the circulation. It plays a key role in maintaining osmolality (the concentration of dissolved particles, such as salts and glucose, in the serum).
Adrenocorticotrophic hormone (ACTH): ACTH is produced by the pituitary gland and its primary function is to stimulate the production and release of cortisol from the adrenal gland cortex.
Adrenaline: Adrenaline is a hormone released from the adrenal glands, together with noradrenaline, its primary role is to prepare the body for fight or flight.
Corticosteroids: Corticosteroids are a class of steroid hormones that are produced in the adrenal cortex of vertebrates. Glucocorticoids are a class of corticosteroids of which cortisol is one. There are also synthetic analogues of these hormones.
Nitric oxide (NO): NO is a key regulator and mediator of numerous processes in the nervous, immune, and cardiovascular systems. For instance, it stimulates vascular smooth muscle relaxation resulting in arterial vasodilation and increasing blood flow.
Heat shock proteins: Heat shock proteins are a family of proteins that are made by cells in response to stress. They chaperone other proteins, guarding them from folding improperly, going astray, or misassembling while forming larger protein aggregates.
HPA axis biology and its dysregulation in disease
HPA axis activity is modulated by CRF and AVP which are secreted from the hypothalamus (Figure 2). These activate the secretion of ACTH from the pituitary which then activates the secretion of the glucocorticoids (or cortisol in humans) from the adrenal cortex (Pariante et al., 2008). Glucocorticoids interact with receptors in various tissues including back at the HPA axis, which is a classic feedback inhibition loop. In this loop, glucocorticoids inhibit both CRF and AVP from the hypothalamus and ACTH from the pituitary. Although still quite poorly understood, cortisol alters the function of the immune system and influences both macrophages and lymphocytes (Padgett, 2003). Both these types of cell are directly responsive to cortisol through the glucocorticoid receptor that they express. This provides a direct link between stress and the immune system.

Figure 2. The influence of stress upon the HPA axis.
The strong anti-inflammatory effects of synthetic glucocorticoids (steroids) in drug therapy are similar to the effects of endogenous glucocorticoids released via the adrenal cortex (Morand, 2001). Defects in HPA axis function are implicated in animal models of rheumatoid arthritis and are hypothesised to contribute towards the disease. Manipulation of HPA axis-driven endogenous anti-inflammatory responses may provide a method for the therapeutic manipulation of inflammatory diseases.
As well as the immune system, glucocorticoids are also known to regulate neuronal survival, neurogenesis, the sizes of complex structures (e.g. the hippocampus), and emotions (Pariante et al., 2008). Reflecting on the HPA axis and its link between stress and the nervous system we are unsurprised to find that it has been found dysregulated in psychiatric disorders, particularly in major or melancholic depression. A significant percentage of depressed patients have increased levels of cortisol in the saliva, plasma, urine, and increased size and activity of the pituitary and adrenal glands. Evidence from clinical studies suggests that childhood trauma is associated with sensitisation of the neuroendocrine stress response, glucocorticoid resistance, increased CRF activity, immune activation, and reduced hippocampal volume (Heim et al., 2008). These neuroendocrine features mirror those found in cases of major depression. The diseases where a dysregulated HPA axis is thought to occur are listed below in Table I.
| Table I: Diseases associated with suppression or activation of immune-mediated inflammation through defects in the HPA axis or its target tissues (adapted from Chrousos, 1995) |
| Suppression of immune meditated inflammatory reactions |
Activation of immune meditated inflammatory reactions |
| |
|
| Increased HPA-axis activity |
Decreased HPA-axis activity |
| Melancholic (major) depression |
Rheumatoid arthritis |
| Chronic stress |
Adrenal insufficiency |
| Chronic alcoholism |
Seasonal or atypical depression |
| |
Hypothyroidism |
| |
Chronic fatigue or fibromyalgia |
| |
|
| |
Resistance to glucocorticoids |
| |
Rheumatoid arthritis |
| |
Steroid resistant asthma |
| |
Systemic lupus erythematosus |
The history of adaptogens
In the late 1940s, scientists of the Far Eastern Division of the Soviet Academy of Sciences in Vladivostok, Siberia, started to study herbs that caused a state of ‘non-specifically increased resistance’ of an organism using animal models and humans (Davydov and Krikorian, 2000). Later, Dr Nikolai Vasilievich Lazarev (1895-1974) a then leading figure in Soviet pharmacology proposed to the scientific community in the mid-1950s that herbs or other substances which could bring about an increased non-specific resistance to stress be called ‘adaptogens’. In the Soviet literature of the time other terminology to describe adaptogenic herbs included, ‘revitalizing therapies’, ‘tonic herbs’, and ‘immunomodulatory substances’. The properties of these adaptogens were outlined in a publication by Brekhman in the 1960s (Brekhman, 1968) as follows:
- The action of an adaptogen should be innocuous and cause minimal disturbance to the normal physiological functions of an organism. It must be absolutely harmless.
- An adaptogenic agent should not be active only in a specific context or against a particular background. It must have a broad therapeutic spectrum of action.
- The action of an adaptogen has to be non-specific, and so resistance to a wide variety of action of harmful factors, whether of a physical, chemical, or biological nature, has to increase.
- An adaptogen has to have a normalising or stabilising action independent of the direction of previous changes.
This somewhat vague definition is not widely known or accepted by mainstream medicine and has been subject to scrutiny for lack of precision (Davydov and Krikorian, 2000). However, the term adaptogen has been refined and defended in work by Alexander Panossian and his colleges (Panossian et al., 2005; Panossian, 2017; Panossian et al., 1999).
Mechanism of action of the adaptogens
It has been proposed, based on animal and human studies, that the action of adaptogens in a single dose is useful in situations that require a rapid response to stress (e.g. heavy exercise) and is related with the modulation by adaptogens of the SAS (Panossian et al., 2005). The SAS provides a rapid response to a stressor and results in increased levels of ATP, NO, cortisol, and various neuropeptides. A characteristic behaviour of adaptogens is that they act as eustressors (or good stressors) and as mild stress mimetics that induce stress-protective responses (Panossian, 2017). Adaptogen treatment can be considered to have a vaccination type effect on the stress response system. For example, in humans, compared with a placebo the adaptogen schisandra increases NO and cortisol in blood plasma and saliva prior to physical exercise, in a similar manner to what normally happens during physical exercise (Panossian et al., 1999). Then, after exercise, in the adaptogen treatment group there are no further increases in NO and cortisol which is related to the enhancement of ordinary physical performance. In addition, it has been observed in C. elegans that adaptogens induce translocation of the DAF-16 transcription factor from the cytoplasm into the nucleus, favouring an increased synthesis of proteins involved in stress resistance and longevity (Wiegant et al., 2009). In this study, it was similarly concluded to the human study, adaptogens are experienced as mild stressors which promote increased stress resistance and, at least in C. elegans, a longer lifespan.
The repeated action of adaptogens on chronic illness is related with the modulation by adaptogens of the HPA axis, which plays a role in the reaction and adaptation to repeated stress (Panossian et al., 2005). To recap, stress triggers secretion of CRF from the hypothalamus, followed by the secretion of ACTH from the pituitary, this stimulates the release of adrenal hormones and NPY to help cope with the stress. Feedback inhibition is initiated by cortisol secreted from the adrenal cortex, which binds to the glucocorticoid receptors (GR) in the brain. This inhibitory signal stops the stress induced release of cortisol and thus cortisol decreases to normal levels in the blood. However, in depression, this feedback inhibition of cortisol via the GR appears blocked and cortisol content in the blood of depressive patients is chronically high (Jovicic et al., 2015). This phenomenon is associated with impaired memory and ability to concentrate, fatigue, and other problems. It is known that the GR are phosphorylated by an enzyme called JNK and this is associated with increased glucocorticoid resistance, increased phosphorylation of GR is observed in patients with depression. Adaptogens stimulate the formation of HSP70 which suppresses elevated JNK and cortisol in the stress response (Panossian, 2017). Thus, adaptogens may help restore a blocked feedback inhibition of the stress response in depressive patients.
Central players modulated by plant adaptogens (such as rhodiola, eleutherococcus, schisandra, ginseng, withania) include the stress hormones cortisol and neuropeptide Y (NPY) and several important mediators of the adaptive stress response (such as NO, stress-activated protein kinases, heat shock proteins (HSP70 and HSP25), and the FOXO transcription factor) (Panossian, 2017). Using experimental models, it has been demonstrated the compromised ability of neurons to express HSP70 is associated with aging-related neurodegeneration (Bobkova et al., 2015). HSP70 treatment has been found to extend lifespan of animals and improve learning and memory, increase curiosity, decrease anxiety, and help preserve synaptic structures that are known to degrade with age. Other studies imply reduced ability to express HSP70 in response to stress may be a common event underlying the aging process (Heydari et al., 1994). Therefore, HSP70 is a candidate pharmacological target of antiaging therapies (Panossian, 2017), although, synthetic chemicals used to induce HSP70 are cytotoxic and are unsafe. However, plant adaptogens have been used safely over a wide dose range (including up to 3000 mg/kg of rat body weight) even with repeated long-term administration (months). The individual plant adaptogens rhodiola, schisandra, eleutherococcus, and their formulation as ADAPT-232, stimulates the expression of HSF-1 and HSP72 in ex vivo neuroglia, provokes HSP72 release from cells, and increases expression of HSP70 in vivo (Panossian et al., 2009; Chiu et al., 2004; Hernández‐Santana et al., 2014; Lee et al., 2009; Li et al., 2014). Of these studies, for example, it was observed continued intake of rhodiola root extract significantly reduced swimming fatigue by increasing glycogen levels, boosted energy generated by lipogenic enzymes, and significantly upregulated HSP70 mRNA (Hernández‐Santana et al., 2014).
Further work on the adaptogen formula, ADAPT-232, has found it promotes expression and release of HSP72 from glioma cells and this requires the action of HSF1 or NPY (Panossian et al., 2012). HSF1 and NPY are likely primary upstream molecular targets of adaptogens in neuroglia. ADAPT-232 appears to act on NPY expression through the upregulation of HSF-1, which lies upstream of HSP72 expression and release. Additionally, ADAPT-232 upregulates both HSP70 and NPY ex vivo (Asea et al., 2013). The activation of NPY by ADAPT-232 promotes HSP70 expression in neuroglia, which helps to maintain homeostasis. It is thought that the induction and release of stress-induced hormone NPY and the stress-induced chaperone HSP70 into circulation is an innate defence response to mild stress and this increases tolerance, adaption, and may also confer longevity (Panossian, 2017). This pathway promotes adaptive and stress-protective effects via varying molecular components throughout the central nervous, sympathetic, endocrine, immune, cardiovascular, and gastrointestinal systems.
Figure 3. More adaptogen herbs. Asian ginseng, rhodiola, and withania (ashwagandha) (from left to right).
It has been demonstrated rhodiola, schisandra, eleutherococcus, withania, and ginseng extend the life span of the nematode C. elegans when stressed (Wiegant et al., 2009; Lee et al., 2007; Kumar et al., 2013), the fruit fly (D. melanogaster) (Jafari, 2007), and the yeast S. cerevisiae (Bayliak et al., 2011). An age-related decline in the ability to induce HSP70 was found in nervous system tissue, (Sherman et al., 2001; Winklhofer et al., 2008) in skeletal and cardiac muscle, and in the liver (Gagliano et al., 2007). It is also noteworthy, inhibition of HSF1 and HSP70 expression occurs in Alzheimer’s disease and is associated with the accumulation of plaques of aggregated β-amyloid peptide (Bhat et al., 2004). Also, it is the heat shock proteins that protect liver cells from the toxic effects of alcohol, heavy metals, xenobiotics, and oxidants. It is known the age-related decline of HSP70 expression contributes to the reduced efficacy of liver detoxification function in aged individuals (Gagliano et al., 2007). A 4-month study in 2-year-old rats showed that, in comparison with a control group, the ADAPT-232 group demonstrated higher liver detoxifying function, better CNS function (memory and learning ability), no development of cardiac insufficiency and hypercholesterolemia, less stress sensitivity, no impaired apoptosis, and no spontaneous tumour promotion (Makarov et al., 2007).
Conclusions
Recent work on the adaptogens in experimental systems has given new insights into the molecular basis on their action on stress response. While no single pathway is responsible for their action, new targets include the heat shock proteins (e.g. HSP70), NPY, and HSF-1. The increase by adaptogens of HSP70 is an interesting discovery since traditionally they have been viewed as life span increasing and this protein is a target of modern longevity research. As discussed in this article, in various experimental (non-human) systems, adaptogens and adaptogenic formula result in increased life expectancy under stressful conditions. It will be interesting to determine whether the same pathways that are involved in this phenomenon in models are also activated in humans with adaptogen treatment. The next part in this article series will be on the adaptogen materia medica and their traditional indications.
For further information regarding the recent scientific developments surrounding the adaptogens, a recent review by Alexander Panossian has been the source of much of the information contained in this article (Panossian, 2017). For more traditional insights into the adaptogenic herbs there is David Winston’s book entitled, ‘Adaptogens: Herbs for Strength, Stamina, and Stress Relief’ (Winston, 2007).
Cautions and contraindications
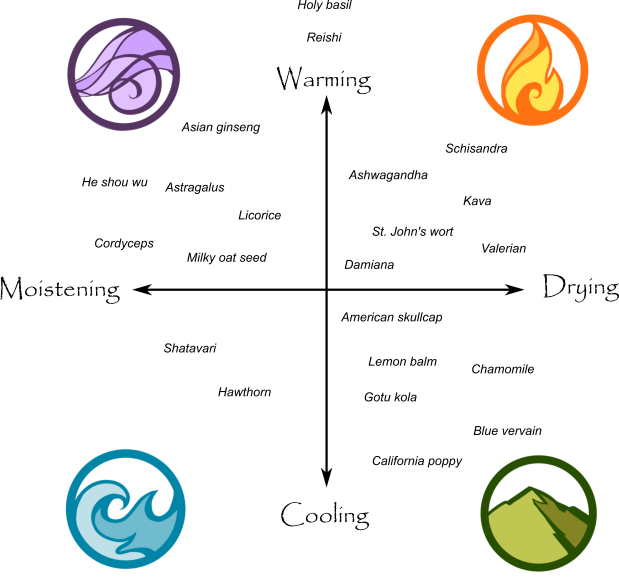
Whilst adaptogens are potentially useful medicines and foods, they can cause insomnia and over stimulation in some individuals (especially the more stimulating ones; e.g. rhodiola, schisandra, Asian ginseng) and depending on them for energy instead of good sleep and eating habits will lead to a ‘sleep debt’ and eventual burnout. It is also sensible to consider the energetic properties of each adaptogen. For instance, schisandra is a drying adaptogen and in a person prone to dryness (e.g. dry skin and slow bowel habits), it may well aggravate this tendency in the longer term. It is best to study the traditional use of the adaptogenic herbs alongside any scientific papers. I recommend reading this PDF by Paul Bergner, an herbalist highly experienced with using the adaptogens if you are thinking of using them on yourself or on others (link).
References
Asea, Alexzander, et al. “Evaluation of molecular chaperons Hsp72 and neuropeptide Y as characteristic markers of adaptogenic activity of plant extracts.” Phytomedicine 20.14 (2013): 1323-1329.
Bayliak, Maria M., and Volodymyr I. Lushchak. “The golden root, Rhodiola rosea, prolongs lifespan but decreases oxidative stress resistance in yeast Saccharomyces cerevisiae.” Phytomedicine 18.14 (2011): 1262-1268.
Bhat, Ratan V., Samantha L. Budd Haeberlein, and Jesús Avila. “Glycogen synthase kinase 3: a drug target for CNS therapies.” Journal of neurochemistry 89.6 (2004): 1313-1317.
Bobkova, Natalia V., et al. “Exogenous Hsp70 delays senescence and improves cognitive function in aging mice.” Proceedings of the National Academy of Sciences 112.52 (2015): 16006-16011.
Brekhman, I. Eleutherokokk (Eleutherococcus). Nauka Publishing House, Leningrad, USSR in Russian. (1968)
Chiu, Po Yee, and Kam Ming Ko. “Schisandrin B protects myocardial ischemia-reperfusion injury partly by inducing Hsp25 and Hsp70 expression in rats.” Molecular and cellular biochemistry 266.1 (2004): 139-144.
Chrousos, George P. “The hypothalamic–pituitary–adrenal axis and immune-mediated inflammation.” New England Journal of Medicine 332.20 (1995): 1351-1363.
Davydov, Marina, and A. D. Krikorian. “Eleutherococcus senticosus (Rupr. & Maxim.) Maxim. (Araliaceae) as an adaptogen: a closer look.” Journal of ethnopharmacology 72.3 (2000): 345-393.
Gagliano, Nicoletta, Fabio Grizzi, and Giorgio Annoni. “Mechanisms of aging and liver functions.” Digestive diseases 25.2 (2007): 118-123.
Heim, Christine, et al. “The link between childhood trauma and depression: insights from HPA axis studies in humans.” Psychoneuroendocrinology 33.6 (2008): 693-710.
Hernández‐Santana, Aaron, et al. “A Rhodiola rosea root extract protects skeletal muscle cells against chemically induced oxidative stress by modulating heat shock protein 70 (HSP70) expression.” Phytotherapy research 28.4 (2014): 623-628.
Heydari, A. R., et al. “Hsp70 and aging.” Cellular and Molecular Life Sciences 50.11 (1994): 1092-1098.
Jafari, Mahtab, et al. “Rhodiola: a promising anti-aging Chinese herb.” Rejuvenation research 10.4 (2007): 587-602.
Jovicic, Milica J., et al. “Modulation of c-Jun N-terminal kinase signaling and specific glucocorticoid receptor phosphorylation in the treatment of major depression.” Medical hypotheses 85.3 (2015): 291-294.
Kumar, Ranjeet, et al. “Withania somnifera root extract extends lifespan of Caenorhabditis elegans.” Annals of neurosciences 20.1 (2013): 13.
Lee, Fang-Tsai, et al. “Chronic Rhodiola rosea extract supplementation enforces exhaustive swimming tolerance.” The American journal of Chinese medicine 37.03 (2009): 557-572.
Lee, Joon-Hee, et al. “Effects of ginsenosides, active ingredients of Panax ginseng, on development, growth, and life span of Caenorhabditis elegans.” Biological and Pharmaceutical Bulletin 30.11 (2007): 2126-2134.
Li, Libo, et al. “Schisandrin B attenuates acetaminophen‐induced hepatic injury through heat‐shock protein 27 and 70 in mice.” Journal of gastroenterology and hepatology 29.3 (2014): 640-647.
Makarov, V. G., et al. “Potential use of plant adaptogen in age related disorders, celebration of the centennial birth of Hans Selye, Budapest, Hungary.” Cell Stress Chaperones 242 (2007).
Morand, Eric F., and Michelle Leech. “Hypothalamic–pituitary–adrenal axis regulation of inflammation in rheumatoid arthritis.” Immunology and cell biology 79.4 (2001): 395-399.
Padgett, David A., and Ronald Glaser. “How stress influences the immune response.” Trends in immunology 24.8 (2003): 444-448.
Panossian, A. G., et al. “Effects of heavy physical exercise and adaptogens on nitric oxide content in human saliva.” Phytomedicine 6.1 (1999): 17-26.
Panossian, Alexander, and H. Wagner. “Stimulating effect of adaptogens: an overview with particular reference to their efficacy following single dose administration.” Phytotherapy Research 19.10 (2005): 819-838.
Panossian, Alexander, et al. “Adaptogens exert a stress-protective effect by modulation of expression of molecular chaperones.” Phytomedicine 16.6 (2009): 617-622.
Panossian, Alexander, et al. “Adaptogens stimulate neuropeptide Y and Hsp72 expression and release in neuroglia cells.” Frontiers in neuroscience 6 (2012).
Panossian, Alexander, et al. “Synergy and antagonism of active constituents of ADAPT-232 on transcriptional level of metabolic regulation of isolated neuroglial cells.” Frontiers in neuroscience 7 (2013).
Panossian, Alexander, Patricia L. Gerbarg, and George Canguilhem. “Potential use of plant adaptogens in age-related disorders.” Complementary and Integrative Therapies for Mental Health and Aging (2015): 197.
Panossian, Alexander. “Understanding adaptogenic activity: specificity of the pharmacological action of adaptogens and other phytochemicals.” Annals of the New York Academy of Sciences (2017).
Pariante, Carmine M., and Stafford L. Lightman. “The HPA axis in major depression: classical theories and new developments.” Trends in neurosciences 31.9 (2008): 464-468.
Sherman, Michael Y., and Alfred L. Goldberg. “Cellular defenses against unfolded proteins: a cell biologist thinks about neurodegenerative diseases.” Neuron 29.1 (2001): 15-32.
Wiegant, F. A. C., et al. “Plant adaptogens increase lifespan and stress resistance in C. elegans.” Biogerontology 10.1 (2009): 27-42.
Winklhofer, Konstanze F., Jörg Tatzelt, and Christian Haass. “The two faces of protein misfolding: gain‐and loss‐of‐function in neurodegenerative diseases.” The EMBO journal 27.2 (2008): 336-349.
Winston, David, and Steven Maimes. Adaptogens: herbs for strength, stamina, and stress relief. Inner Traditions/Bear & Co, 2007.


























_wikicommons")